
California Partners in Flight Desert Bird Conservation Plan
Verdin (Auriparus flaviceps )
Prepared by: Roy T. Churchwell (roychurchwell@hotmail.com)
San Francisco Bay Bird Observatory
1290 Hope Street
Alviso, CA 95002
RECOMMENDED CITATION
Churchwell, R.T. 2007 Verdin (Auriparus flaviceps). In The Desert Bird Conservation Plan. California Partners In Flight. http://www.prbo.org/calpif/htmldocs/desert.html
SHORTCUTS
SUBSPECIES STATUS
Within the Desert Plan’s extent, A. f. acaciarum from southeast California, southern Nevada, southern Utah, Arizona, Baja California Norte to Ensenada, central Sonora. Nominate A. f. flaviceps central Baja and western Sonora (Webster 1999).
Management Status
Partners in Flight Stewardship Species (Global population in a single biome)
DISTRIBUTION
The Verdin is a resident species of the Southwestern desert including the Mojave, Sonora, and Chihuahuan Deserts (Webster 1999). This species range includes southeastern California (Grinnell and Miller 1944, Webster 1999), the southern tip of Nevada (Johnson and Richardson 1952) to the extreme southwestern corner of Utah (Behle et al. 1963, Behle 1976), it then extends through Arizona excluding the northeast corner of the state (Webster 1999, Wise-Gervais 2005), includes the southwest corner of New Mexico, and includes the western half of Texas (Griscom and Crosby 1925) up to, but excluding the Texas Panhandle (Webster 1999) and including the extreme southwestern corner of Oklahoma (Baumgartner and Baumgartner 1992, Webster 1999). In Mexico the range includes most of the Baja Peninsula except the northwest corner, most of the state of Sonora and the northwest corner of Sinaloa, also all but the western edge of Chihuahua, Durango, Zacatecas, the northeastern edge of Jalisco, and the northern edge of Michoacán and México. Then the range extends north to the eastern border of Neuvo León and finally east to the coast including the northern portion of Tamaulipas (Webster and Orr 1954, Webster 1999, Howell and Webb 1995).
The species was once found in coastal San Diego County, the Tijuana Valley, and parts of Baja California where it is now absent (Kading 1905, Garrett and Dunn 1981, Harrap and Quinn 1995). Verdin were also found around Furnace Creek in Death Valley (Grinnell 1931), but have been absent after declining in the early 1970s (Garrett and Dunn 1981 Small 1994). On the eastern edge of its range it is thought to have expanded in Arizona (Lloyd et al. 1998) and also from central Texas up to Oklahoma, although nesting has not been reported in Oklahoma since 1977 (Carnes 2004, Baumgartner and Baumgartner 1992, Carnes 2004). In Mexico it may have recently expanded its range into those areas of Zacatecas, Durango, and Chihuahua where it now occurs (Leopold 1959, Webster 1978, in Webster 1999).
The first type specimen for Verdin was collected by a Russian Naturalist I.G. Vosnesensky around 1841 along the coast near Loreto, Mexico (Grinnell 1931). Now there are seven recognized subspecies, of which only one occurs in California (Auriparus flaviceps acaciarum). The eastern extent of the range of this subspecies has not been agreed upon, but it extends at least to the Colorado River (Hellmayr 1934 and Snow 1967 in Webster 1999) and may occur across Arizona (Phillips 1986 in Webster 1999).
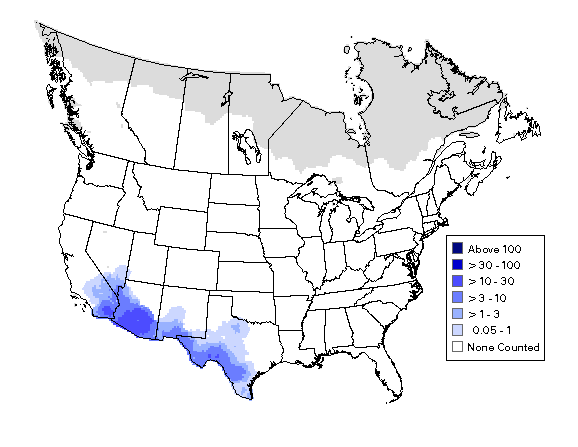
Figure 1. Verdin distribution and density in the U.S., ( http://www.mbr-pwrc.usgs.gov/bbs/htm03/ra2003_blue/ra07460.htm)
Distribution in the West Mojave Planning Area:
Auriparus flaviceps acaciarum is the single subspecies representative of Verdin in the West Mohave Planning Area. Using Christmas Bird Count data, Root (1988) determined that the Mojave Desert has the greatest densities of Verdin at 4.52 individuals/ hour. The species occupies thorn scrub desert habitat of the southeastern portion of the state including the Colorado River Valley (Daggett 1902, Hollister 1908, Stephens 1903, Webster 1999). The species range extends west from the Colorado River including all of Imperial County and the very eastern portion of San Diego County including Anza Borego (Rossem 1911, McCaskie and Banks 1964, Small 1994). The range extends across half of Riverside County to Palm Springs (Grinnell 1904, Miller 1932, Small 1994), and across San Bernardino County (Grinnell 1901, Mailliard and Grinnell 1905, Carter 1937) to the northeast corner of Los Angeles County and the southeast corner of Kern County, but excluding the south- and northwestern corners of San Bernardino county. Populations in Los Angeles and Kern Counties are local; in Los Angeles County at Saddleback Butte State Park and near Lancaster, CA and in Kern County in the area of Rosamond and Ancient Valley near Willow Springs, CA (Small 1994). Finally northern extent of the range is in the southeastern corner of Inyo County and locally along the Amaragosa River (Grinnell 1934, Wauer 1962) and near springs in the Panamint Mountains (Wauer 1964). Their former range included the Tijuana River Valley in San Diego County (Kaeding 1905), and the intermittent population at Furnace Creek, Death Valley National Monument (Grinnell 1934, Wauer 1962). Vagrants have been found in Chula Vista and San Elijo Lagoon San Diego County and Ming Lake near Bakersfield, Kern County, CA (Small 1994).
NATURAL HISTORY
The Verdin is a small insectivorous passerine of 6 to 8 grams and 9 to 11 cm in length (Webster 1999). Although systematics of the species is in question, the species is currently thought to be the only North American member of the Remizidae family as determined by behavior, calls, and DNA-DNA hybridization (Taylor 1967, Cramp and Perrins 1993, Sheldon and Gill 1996). Due to the bright yellow facial pattern of this species, the adults are not easily confused with other species. Young birds (April to August) lacking this character can easily be confused with other species like the Juniper or Oak Titmice, Bushtit, Lucy’s Warbler, and Bell’s Vireo (Bent 1946, Sibley 2003). Males can sometimes be distinguished from females by a brighter yellow on the face. In the hand, the yellow of the female extends 6-15 mm below the bill and the male 12-23 mm below the bill. Also the female wing length ranges 48-54 mm while the male ranges 50-57 mm (Pyle 1997).
The Verdin is a resident species, and is not thought to be a migrant (Bent 1946). Young birds have been documented to disperse 0.5 to 3 km from their natal territory (Taylor 1967). There is some evidence that birds may disperse in the winter (Taylor 1967, Harrap and Quinn 1995), but that both males and females are territorial in the winter (Harrap and Quinn 1995). These wintering birds often make up the breeding population of that area the next summer (Taylor 1971). The Verdin may join foraging flocks while they feed in the territory, but leave the group as it passes on to other foraging areas (Austin and Smith 1972).
The Verdin is predominately insectivorous (Bent 1946, Taylor 1971) generally feeding in the outer branches of spiny shrubs (Parker 1986) like paloverde (Cercidium spp.), ironwood (Olneya tesota), and Acacia spp. They also feed in riparian habitat (Taylor 1971), and are one of the few species to be found commonly feeding in upland creosote (Larrea tridentate) (Anderson and Anderson 1946). Parker (1986) found that Verdin spent over half of their non-flight time in spiny shrubs that only composed 27% of the vegetative cover in an Arizona wash habitat. In the wash habitat of Arizona Verdin divided their time between feeding in the washes and the adjacent upland flats (Parker 1986). Verdin are known to feed on flowers (Pickens 1929) piercing the base of the tube and drinking the nectar (Webster 1999), visiting saguaro (Carnegiea gigantea) flowers (Taylor 1971), or pulling the flowers off creosote bush and then dropping them (Parker 1986). They also feed on berries and bring them to the nest for their young, and have been observed feeding on palm (Phoenix spp.) dates in the fall (Taylor 1971).
Unlike many species the verdin’s nest is often obviously placed in the outer branches of a spiny shrub including paloverde, acacia, mesquite (Prosopis spp.), cholla cactus (Cylindropuntia spp.), (Bent 1946, Taylor 1971) or ironwood (Jaeger 1949). Other nests are found less frequently in non-thorn bearing shrubs including Tamarisk spp., mistletoe (Phoradendron californicum), almond (Prunus dulcis), elm (Ulmus spp.), Gambel’s oak (Quercus gambelii), lemon, and willow (Salix spp.) to name a few (Taylor 1971). Many descriptions have been made of the Verdin nest (Gilman 1902, Bailey 1923, Hensley 1954, Bent 1946, Taylor 1971, Austin 1976, Webster 1999). The nest is normally spherical or elongated and 18.4 x 13.6 cm. The inside cavity is 4-6 cm in diameter and the entrance hole is 2.5 cm in diameter (Webster 1999). Taylor’s (1971) description is very detailed, describing the three layers of the nest including the twig outside frame including the thorn guarded entrance, the middle layer of small twigs, leaves, spider webbing, and moss, and the nest cup of feathers, fur, or wool. The Verdin constructs both breeding and roosting nests. Two kinds of roosting nests have been found: small lighter nests built in the late summer and larger nests resembling breeding nests in the fall and winter (Taylor 1971). Breeding nests are started by the male and then the female helps complete the structure. While the female is brooding the eggs, she will continue to bring nest material to the nest (Taylor 1971, Webster 1999). Taylor (1971) did not find the nest entrance of nests in his study to be arranged in any direction. Austin (1976) on the other hand found that spring nests were oriented away from the prevailing wind direction while late summer nests were oriented towards the prevailing wind direction. Winter nests in another study were randomly oriented (Buttemer et al. 1987). Verdin nests are very sturdy and persist for many years after they are built giving the illusion that there are more Verdins in the area than there are (Bent 1946). Nests from past years are sometimes used by other nesting birds including Lucy’s Warbler (Swarth 1905), Black-tailed Gnatcatcher (Finch 1982), Bewick’s Wren, and Cactus Wren (Taylor 1971).
The nest contains 3 - 6 but usually four eggs (Baicich and Harrison 1997, Webster 1999). The young are born altricial after an incubation period of 14 - 18 days (Taylor 1971, Baicich and Harrison 1997). The nestlings are first fed by the female, and although originally reported, the young are not fed a regurgitated substance in the beginning (Wheelock 1920, Taylor 1971, Webster 1999). After 5 - 7 days the male helps to feed the young (Webster 1999). The young leave the nest 17 - 21 days after hatching (Baicich and Harrison 1997), but return to the breeding nest to roost for up to 2 weeks (Webster 1999). The male broods the young at night after fledging, as the female will lay a new clutch as soon as 2 days after fledging (Webster 1999). Males and females do not roost together, but both sexes can be found roosting with young (Taylor 1971). A Verdin pair will attempt to complete two successful clutches (Taylor 1971, Webster 1999). There are a few observations of cowbird parasitism of Verdin nests although it is not common (Friedmann 1934, Taylor 1971, Webster 1999), and one published instance of Verdins feeding cowbird young (Austin 1970). The nest entrance may need to be modified in order to make it possible for the cowbird to occupy the nest (Friedmann 1925, Bent 1946). There is one observation of a Verdin laying its egg in a northern mocking bird nest, but the nest was taken by a predator before it could be determined if it would hatch (Carter 1987).
The Verdin is a small bird living in a habitat with extreme seasonal differences in temperature, and this species has been the focus of a great deal of research on how the species copes with these extremes. Goldstein (1974) found the Verdin to be physiologically adapted to living in a cold climate because the species had a higher than expected standard metabolic rate and lower than expected thermal conductance value. By roosting in an insulated nest, the Verdin may save as much as 29-49% in energy when compared to a bird of equal size roosting in the open air (Buttemer et al. 1987, Webster 1989). Another species has been observed using the insulating qualities of the Verdin nest, when as many as 15 black-tailed gnatcatchers were found roosting in a Verdin nest for several nights (Walsberg 1990). Taylor (1978) found that Verdins spent less time moving when temperatures increased to greater than 32ºC. Also, during the warmest parts of the day, it has been found that Verdin use shaded areas as refugia, saving as much as 5 times the evaporative water loss than if they were in the sun (Wolf et al. 1996).
HABITAT REQUIREMENTS
The Verdin is a bird of the desert scrub mostly found in desert riparian areas and desert washes. In California the species is normally associated with paloverde, mesquite, catclaw, creosote bush, smoke tree. They are less commonly found in areas occupied by Joshua tree. They also once occurred in the seep willow and tree tobacco of the Tijuana River Valley (Garrett and Dunn 1981). The species is normally found at low elevations, but has been found up to 915 m in California (Harrap and Quinn 1995).
Although this species is often found near water, it does not need water to survive and has been found as many as 3 miles away from the nearest water supply (Bent 1946).
In creosote desert in Arizona, Tomoff (1974) found Verdins were denser in areas with greater foliage density when compared to areas with lower foliage density. In Tucson where the effect of urbanization on the bird community has been documented, most of the research indicates that Verdin benefit from urbanization to a point due to increased vegetation compared to their native habitat (Emlen 1974, Mills et al. 1989.) In Tucson, Germaine et al. (1998) found Verdin were significantly negatively associated with house density, paved areas, urban open exotic plant areas, distance to vegetation patch, and distance to riparian and were positively associated with upland Sonoran vegetation and undisturbed wash. In Tempe, Verdin were found to be more common in urban areas than in natural habitat (Resenberg et al. 1987)
POPULATION STATUS
The Verdin is a common species of the desert washes of the Mojave Desert. Root (1988) found the highest densities of the Verdin in the Mojave Desert. The Verdin is not listed, except as a Partners in Flight Stewardship Species because its global population is found in a single biome (Rich et al. 2004).
The Breeding Bird Survey (BBS) reports that in the Sonoran Desert between 1966 and 2007 the Verdin shows a significant trend of decline (-3.3% annually, P = 0.04). Populations in the Mojave Desert show an even faster rate of decline (-5.4%), but this result was not significant at the α = 0.05 threshold (P = 0.17) – perhaps due to a lower sample size in the Mojave Desert (Sauer et al. 2008). The BBS reports a relative abundance of 3.56 birds/route (Sauer et al. 2005). The Christmas Bird Count reports between 1959 and 1988, that survey-wide the Verdin has a stable population with a relative abundance of 2.09 birds/100 party hours. The survey does not report numbers for California, and although it may not be applicable to California, in Arizona the species shows a significantly increasing trend and a relative abundance of 5.04 birds/100 party hours (Sauer et al. 1996). Webster (1999) reports species density from several studies in California (16-28 individuals/ 40 ha in the Colorado Desert, 20-24 individuals/ 40 ha in the Lower Colorado River Valley, and 17.4 individuals/ 40 ha in Riverside County) for the Verdin Bird of North America account.
The species has a stable population over most of its range in the Mojave Desert, but the decline of the species has been documented in two satellite populations in the Tijuana Valley (Kaeding 1905, Small 1994) and at Furnace Creek in Death Valley (Grinnell 1934, Wauer 1962).
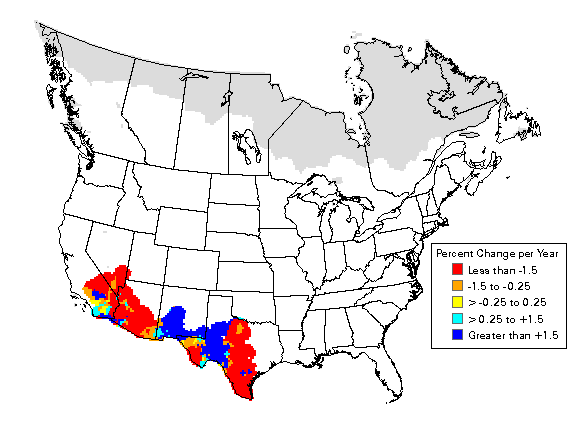
Figure 2. Verdin population trend, 1966-2003. http://www.mbr-pwrc.usgs.gov/bbs/htm03/trn2003/tr07460.htm
THREATS ANALYSIS
The Verdin is one desert species that seems to adjust to minimal habitat disturbance. Patten and Rotenberry (1998) found the Verdin population was stable at their study site after much of the riparian vegetation was removed from a portion of their study area. Also, several authors found that Verdin were successfully established in suburban areas, but that their numbers decline with the amount of development (Rosenberg et al. 1987, Germaine et al. 1998).
Verdin are also found to inhabit riparian areas invaded by salt cedar along the Colorado River (Hunter et al. 1988), but the authors thought the birds may not inhabit the same habitat along the middle Rio Grande because of colder winters in the Rio Grande area. There is the potential for salt-cedar to impact Verdin density in this habitat compared to native habitats, but there was no information found on how salt cedar influences the density of this species.
There is one study documenting the impact of grazing on a riparian bird community along the San Pedro River in Arizona of which Verdin was one of the species (Krueper et al. 2003). In this study, the Verdin population declined with grazing but the trend was not statistically significant. Also, Brook (1999) found Verdin density increased 200% in fenced areas in the Desert Tortoise Research Natural Area that prevented grazing and off-highway vehicle use in the area.
In the Mojave Desert region the Verdin is most dependent on arid riparian wash habitat. These areas are increasingly used for recreational purposes including off-highway vehicle use and firewood gathering. No published information was found on impacts from recreation in desert washes, although PRBO Conservation Science is currently working on these questions.
One final question that is still standing is the possible impact of global warming on the Verdin. Most models indicate the desert Southwest will become warmer as global climate change sets in. For a species like the Verdin that spends the hottest parts of the day in cooler shaded areas, warmer temperatures could have a significant impact on their daily time budget. For example these refuge areas may become less available, or the Verdin may be required to spend even more of their time in these refuge areas leaving less time for other time budget tasks (Wolf 2000).
BIOLOGICAL STANDARDS
The Verdin is most dependent on the thorn scrub of the desert riparian and wash habitat of the Mojave Desert. As this habitat becomes more and more popular as a recreation area a need may arise for regulating recreational activity in portions of this desert habitat to protect the Verdin and other species dependent on thorn scrub habitats.
There is evidence that the Verdin can inhabit areas with salt cedar, but further research could elucidate the influence of salt-cedar invasion on Verdin abundance, survival, and fecundity.
There are several satellite Verdin populations in California, including those located in Los Angeles and Kern Counties and the Panamint Mountains. These populations should be monitored to better understand population regulation of Verdin, and determine if the population is increasing, decreasing, or both depending on the location. Indications from Furnace Creek and the Tijuana Valley suggest that Verdin distribution may be shrinking in California.
Lastly, as the impacts of global climate change are felt, the Verdin population along with populations of other desert species should be monitored. Although the final outcome of global climate change is still in question, any change in the desert climate could have the potential to greatly impact survival of desert birds. With the extensive background information on Verdin physiology and the response of this species to heat stress and other environmental factors, the Verdin is a prime candidate for research on impacts of a changing climate.
Anderson, A.H. and A. Anderson. 1946. Notes on the use of the creosote bush by birds. Condor 48:179.
Austin, G.T. 1970. Breeding birds of desert riparian habitat in southern Nevada. The Condor 72:431-436.
Austin, G.T. and E.L. Smith. 1972. Winter foraging ecology of mixed insectivorous bird flocks in oak woodland in Southern Arizona. The Condor 74:17-24.
Austin, G.T. 1976. Behavior adaptations of the Verdin to the desert. The Auk 93:245-262.
Baicich, P.J. and C.J.O. Harrison. 1997. A guide to the nests, eggs, and nestlings of North American Birds, Second edition. Academic Press, San Diego, California.
Bailey F.M. 1923. Fifteen Arizona Verdins’ nests. The Condor 25:20-21.
Baumgartner, F.M. and A.M. Baumgartner. 1992. Oklahoma Bird Life. University of Oklahoma Press, Norman, Oklahoma.
Behle, W.H., J.B. Bushman, and C.M. White. 1963. Distributional data on uncommon birds in Utah and adjacent states. Wilson Bulletin 75:450-456.
Behle, W.H. 1976. Mohave Desert avifauna in the Virgin River Valley of Utah, Nevada, and Arizona. The Condor 78: 40-48.
Bent, A.C. 1946. Life histories of North American Jays, Crows, and Titmice, Part two. Pp. 427-438. Dover Publications Inc., New York, New York.
Brook, M. 1999. Effects of protective fencing on birds, lizards, and black-tailed hares in the Western Mojave Desert. Environmental Management 23:387-400.
Buttemer, W.A., L.B. Astheimer, W.W. Weathers, A.M. Hayworth. 1987. Energy savings attending winter-nest use by Verdins (Auriparis flaviceps). The Auk 104:531-535.
Carnes, G. 2004. Verdin (Auriparus flaviceps). In Oklahoma Breeding Bird Atlas (Reinking, D. ed). Pp. 302-303. University of Oklahoma Press, Norman, Oklahoma.
Carter, F. 1937. Bird life at Twentynine Palms. The Condor 39:210-219.
Carter, M.D. 1987. An incident of brood parasitism by the Verdin. The Wilson Bulletin 99:136.
Cramp, S. and C.M. Perrins, eds. 1993. Remizidae. Pp. 376-396 In The birds of the western Palearctic. Vol. VII. Oxford University Press, Oxford, U.K.
Daggett, F.S. 1902. Winter observations on the Colorado Desert. The Condor 4: 37-39.
Emlen, J.T. 1974. An urban bird community in Tucson, Arizona: derivation, structure, regulation. The Condor 76:184-197.
Finch, D.M. 1982. Interspecific nest use by aridland birds. The Wilson Bulletin 94:582-584.
Friedmann, H. 1925. Notes on the birds observed in the Lower Rio Grande Valley of Texas during May, 1924. The Auk 42:537-554.
Friedmann, H. 1934. Further addition to the list of birds victimized by the cowbird. The Wilson Bulletin 46:25-36.
Garrett, K. and J. Dunn. 1981. Birds of Southern California: status and distribution. Artisian Press, Los Angeles, California.
Gillman, M.F. 1902. Notes on the Verdin. Condor 4:88-89.
Germaine, S.S., S.S. Rosenstock, R.E. Schweinsberg, W.S. Richardson. 1998. Relationship among breeding birds, habitat, and residential development in greater Tucson, Arizona. Ecological Applications 8:680-691.
Goldstein, R.B. 1974. Relation of metabolism to ambient temperature in the Verdin. Condor 76:116-119.
Grinnell, J. 1901. Midwinter birds at Barstow. The Condor 3:70-71.
Grinnell, J. 1904. Midwinter birds at Palm Springs, California. The Condor 6:40-45.
Grinnell, J. 1934. Further observations upon the bird life of Death Valley. The Condor 36:67-72.
Grinnell, J. and A.H. Miller. 1944. The distribution of California Birds. Cooper Ornithological Club 27:310.
Griscom, L. and M.S. Crosby. 1925. Birds of the Brownsville region southern Texas. The Auk 42:432-440.
Harrap, S. and D. Quinn. 1995. Chickadees, tits, nuthatches, and treecreepers. Pp. 234-237. Princeton University Press, Princeton, New Jersey.
Hensley M. 1954. Ecological relations of the breeding bird population of the desert biome in Arizona. Ecological Monographs 24:185-208.
Hensley M.M. 1959. Notes on the nesting of selected species of birds of the Sonoran Desert. Wilson Bulletin 71:86-92.
Hollister, N. 1908. Birds of the region about Needles, California. The Auk 25:455-462.
Howell, S.N.G. and S. Webb. 1995. A Guide to the Birds of Mexico and Northern Central America. P. 551. Oxford University Press, Inc. New York, New York.
Jaeger, E.C. 1949. Relation of the desert ironwood tree to nesting. The Auk 66:211-212.
Johnson, N.K., and F. Richardson. 1952. Supplementary bird records for Nevada. The Condor. 54:358-359.
Kading, H.B. 1905. Birds from the west coast of lower California and adjacent islands. The Condor 7:134-138.
Krueper, D., J. Bart, and T.D. Rich. 2003. Response of vegetation and breeding birds to the removal of cattle on the San Pedro River, Arizona (U.S.A.). Conservation Biology 17:607-615.
Leopold, A.S. 1959. Wildlife of Mexico. University of California Press, Berkeley, California.
Lloyd, J., R.W. Mannan, S. Destefano, and C. Kirkpatrick. 1998. The effects of a mesquite invasion on a southern Arizona grassland bird community. The Wilson Bulletin 110:403-408.
Mailliard, J. and J. Grinnell. 1904. Midwinter birds on the Mojave Desert. The Condor 7:101-102.
McCaskie, R.G. and R.C. Banks. 1964. Occurrence and migration of certain birds in southwestern California. The Auk 81:353-361.
Miller, L. 1932. The Saw-whet Owl in the desert. The Condor 34:258.
Mills, G.S., J.B. Dunning, Jr., and J.M Bates. 1989. Effects of urbanization on breeding bird community structure in southwestern desert habitats. The Condor 91:416-428.
Hunter, W.C., R.D. Ohmart and B.W. Anderson. 1988. Use of exotic salt cedar (Tamarix chinensis) by birds in arid riparian systems. The Condor 90:113-123.
Patten, M.A. and J.T. Rotenberry. 1998. Post-disturbance changes in a desert breeding bird community. The Journal of Field Ornithology 69:614-625.
Parker, K.C. 1986. Partitioning of foraging space and nesting sites in a desert shrubland bird community. American Midland Naturalist 115:255-267.
Pickens, A.L. 1929. Bird pollination problems in California. Condor 31:229-232.
Pyle, P. 1997. Identification guide to North American birds. Pp. 347-348. Slate Creek Press, Bolinas, California.
Rich, T.D., C.J. Beardmore, H. Berlanga, P.J. Blancher, M.S.W. Bradstreet, B.S. Butcher, D.W. Demarest, E.H. Dunn, W.C. Hunter, E.E. Inigo-Elias, J.A. Kennedy, A.M. Martell, A.O. Panjabi, D.N. Pahley, K.V.
Rosenberg, C.M. Rustay, J.S. Wendt, T.C. Will. 2004. Partners in flight North American landbird conservation plan. Cornell Lab of Ornithology, Ithaca, New York.
Root, T. 1988. Atlas of Wintering North American Birds: An Analysis of Christmas Bird Count Data. P. 172. The University of Chicago Press, Chicago, Illinois.
Rosenberg, K.V., S.B. Terrill, and G.H. Rosenberg. 1987. Value of suburban habitats to desert riparian birds. The Wilson Bulletin 99:642-654.
Rossem, A.V. 1911. Winter Birds of the Salton Sea Region. The Condor 13:129-137.
Sheldon, F.H. and F.B. Gill. 1996. A reconsideration of songbird phylogeny, with an emphasis on the evolution of titmice and their Sylvioid relatives. Systematic Biology 45:473-495.
Sibley, D.A. 2003. The Sibley field guide to birds of Western North America, P. 323 Alfred A. Knopf, Inc. New York, New York.
Small, A. 1994. California Birds: Their Status and Distribution. P. 180. Ibis Publishing Company, Vista, California.
Stephens, F. 1904. Birds notes from eastern California and western Arizona. The Condor 5:100-105.
Sauer, J. R., S. Schwartz, and B. Hoover. 1996. The Christmas Bird Count Home Page. Version 95.1. Patuxent Wildlife Research Center, Laurel, MD. http://www.mbr-pwrc.usgs.gov/bbs/cbc.html. February 19, 2007.
Sauer, J. R., J. E. Hines, and J. Fallon. 2008. The North American Breeding Bird Survey, Results and Analysis 1966 - 2008. Version 5.15.2008. USGS Patuxent Wildlife Research Center, Laurel, MD. http://www.mbr-pwrc.usgs.gov/bbs/
Swarth, H.S. 1905. Summer birds of the Papago Indian Reservation and of the Santa Rita Mountains, Arizona. The Condor 7:47-50.
Taylor, W.K. 1971. A breeding biology study of the Verdin, Auriparus flaviceps in Arizona. American Midland Naturalist 85:289-328.
Taylor, W.K. 1978. Daily time budget of the post nesting verdin. The Auk 95:247-251.
Tomoff, C.S. 1974. Avian species diversity in desert scrub. Ecology 55:396-403.
Walsberg, G.E. 1990. Communal roosting in a very small bird: consequences for the thermal and respiratory gas environments. The Condor 92:795-798.
Wauer, R.H. 1962. A survey of the birds of Death Valley. The Condor 64:220-233.
Wauer, R.H. 1964. Ecological distribution of the birds of the Panamint Mountains, California. The Condor 66:287-301.
Webster, J.D., and R.T. Orr. 1954. Miscellaneous notes on Mexican Birds. The Wilson Bulletin 66: 267-269.
Webster, J.D. 1978. The avifauna of the southern part of the Chihuahuan Desert. Pp. 559-566 In Transactions of the Symposium on the Biological Resources of the Chihuahuan Desert Region (Wauer, R.H., and D.H. Ruskind, eds.). U.S. Department of Interior, National Park Service Transactions and Procedures Series No. 3.
Webster, M.D. 1989. Overnight weight loss by wintering verdins. The Condor 91:983-985.
Webster, M.D. 1999. Verdin (Auriparusl flaviceps). In The Birds of North America, No. 470 (A. Poole and F. Gill, eds.). The Birds of North America, Inc., Philadelphia, Pennsylvania.
Wheelock, I.G. 1920. Birds of California, 5th edition, P. 578. A.C. McClurg and Company, Chicago, Illinois.
Wise-Gervais, C. 2007. Verdin (Auriparus flaviceps). Pp. 390-391. In Arizona Breeding Bird Atlas. (Corman, T. and C. Wise-Gervais, eds.). University of New Mexico Press, Albuquerque, New Mexico.
Wolf, B.O., K.M. Wooden, and G.E. Walsberg. 1996. The use of thermal refugia by two small desert birds. The Condor 98:424-428.
Wolf, B.O. 2000. Global warming and avian occupancy of hot deserts; a physical and behavioral perspective. Revista Chilena de History Natural v 73. http://www.scielo.cl/scielo.php?script=sci_arttext&pid=S0716-078X2000000300003&lng=en&nrm=iso/&tlng=en (Feb. 25, 2007)